Alvi Milliana1,
A S Noorhamdani2,
Sri Poeranto3,
Kusworini Handono4,
Sumarno Reto Prawiro2 ![]() ,
Avin Ainur Fitrianingsih1,
Lailia Nur Rachma1
,
Avin Ainur Fitrianingsih1,
Lailia Nur Rachma1
For correspondence:- Sumarno Prawiro Email: retoprawiros@yahoo.com Tel:+81555840099
Received: 15 September 2016 Accepted: 20 January 2017 Published: 25 February 2017
Citation: Milliana A, Noorhamdani AS, Poeranto S, Handono K, Prawiro SR, Fitrianingsih AA, et al. Antibodies against Shigella flexneri adhesion molecule outer membrane protein (OMP) can cross-react with OMPs of some Shigella species. Trop J Pharm Res 2017; 16(2):255-261 doi: 10.4314/tjpr.v16i2.1
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To identify a Shigella flexneri hemagglutinin outer membrane protein (OMP) as an adhesion factor and examine its ability to cross-react with the OMPs of other Shigella species.
Methods: OMP was isolated from the bacterium S. flexneri after shaving the pili using a pili bacterial cutter in a solution of 0.5 % n-octyl-β-D-glucopyranoside. Hemagglutination was evaluated using mice erythrocytes. The molecular weight (MW) of the OMPs was profiled by SDS-PAGE. Adhesion molecules were identified by calculating the index of adhesion. Thereafter, antigen cross-reaction with antibodies was examined by Western blotting,
Results: The S. flexneri adhesion molecule OMP has a MW of 28 kDa. An antibody against the 28 kDa S. flexneri OMP adhesion molecule cross-reacted with different MW OMPs from other Shigella species.
Conclusion: This study showed that the 28 kDa S. flexneri OMP is a hemagglutinin adhesion protein, and antibodies against it can cross-react with the OMPs of other Shigella species.
Introduction
Diarrhea is the fourth leading cause of morbidity and mortality worldwide, and Shigella species cause an estimated 164.7 million cases of diarrhea annually. In developing countries, the 163.2 million cases resulted in 1.2 million deaths [1]. In South Jakarta, the predominant isolate was S. flexneri, which is often isolated in other developing regions, such as Bangladesh, Pakistan, and Africa [2]. S. flexneri is endemic in developing countries [1,3].
Shigella is a highly infectious pathogen, because only 10 – 100 ingested bacteria are sufficient to cause disease [4]. The success of a pathogenic bacterial infection requires attachment to host cells, tissue colonization, and cell invasion [5].
The initial stage of infection is adhesion between the bacteria and host cells. Outer membrane proteins (OMPs) include proteins that mediate the attachment of bacteria to the host cell more closely than do pili [6,7]. A 49.8 kDa Shigella dysenteriae hemagglutinin pili protein that serves as an adhesion protein was found previously [8]. Oral immunization with this 49.8 kDa S. dysenteriae protein can increase intestinal secretory IgA (s-IgA) levels, inhibit bacterial colonization in the colon, and prevent damage to colon epithelial cells [9]. There are two S. dysenteriae pili protein subunits, of 7.9 and 49.8 kDa. The S. dysenteriae protein prevented diarrhea in a mouse ligated ileal loop (MLIL) model [10].
The World Health Organization (WHO) has prioritized the development of safe, effective vaccines for shigellosis, and several candidate vaccines are being developed currently [1,11]. OMPs have been used successfully as vaccine antigens to prevent Lyme disease and pertussis based on their roles as adhesion molecules [11-13]. The ideal Shigella vaccine should be able to induce an immune response against all Shigella spp. [11].
Methods
Sample preparation
The bacteria Shigella flexneri 4a, Shigella dysenteriae 2a, Shigella boydii serotype 10, and Shigella sonnei were obtained from the Yogyakarta Regional Health Laboratory. Culture isolates of the Shigella species were grown on MacConkey medium at 37 °C for 24 h and then transferred to Petri dishes containing Salmonella-Shigella Agar (SSA) medium at 37 °C for 24 h. The resulting bacterial cultures were dissolved in 10 mL phosphate-buffered saline (PBS) (pH 7.4), added to bottles containing 500 mL Brain Heart Infusion (BHI), and shaken in a water bath for 30 min at 37 °C.
Isolation of OMP
The Shigella OMP was isolated in two stages. The first stage involved cutting the bacterial cell pili, and the second involved isolating the OMP from the bacterial cell after pili cutting, as described in our previous study [14]. The OMP was isolated according to the method of Evans and Evans [15]. The pellet from the last centrifugation step was resuspended in PBS (pH 7.4) until the volume reached 15-fold, and then n-octyl-β-d-glucopyranoside was added at concentrations up to 0.5 %. The resulting solution was homogenized using a vortex and centrifuged at 12,000 rpm at 4 °C for 15 min. This treatment was repeated three times to give a supernatant rich in OMP.
Purification of Shigella spp. OMP
The molecular weight of the OMP was determined using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) [16]. The OMP was purified by electroelution [12]. The elution product was dialyzed for 24 h in a glass beaker with sterile PBS and stirred at 4 °C for 24 h. The obtained protein was precipitated by centrifugation at 12,000 rpm for 15 min at 4 °C.
Hemagglutination test
The hemagglutination test was performed according to the method of Hanne and Finkelstein with modifications [14]. First, 50 µL PBS were placed in the wells of a Microplate V, and 50 µL serial dilutions of the sample were added. Then, 50 µL 0.5 % mice erythrocyte suspension were added to each well, and the plate was shaken gently for 1 min on a plate rotator. The hemagglutination titer was the lowest dilution causing agglutination of the red blood cells.
Isolation of mouse enterocytes
Mice enterocytes were isolated using the Weisser method [17]. The mice were anesthetized using chloroform, their stomachs opened, and the small intestines removed. The small intestine was cut into small pieces and washed several times until clean using a solution of PBS containing 1 mM dithiothreitol (DTT) at 4 °C. Then, the intestinal tissue was added to a solution containing 1.5 mM KCl, 9.6 mM NaCl, 27 mM Na-citrate, 8 mM KH2PO4, and 5.6 mM Na2HPO4 at pH 7.4. The solution was incubated on a shaking incubator at 37 °C for 15 min. The supernatant was discarded and the tissue was transferred to a solution containing 1.5 mM EDTA and 0.5 mM DTT. The liquid was shaken vigorously for 15 min at 37 °C, and the supernatant was discarded. The tissue was washed with PBS and then centrifuged at 1500 rpm at 4 °C for 5 min; this was repeated at least three times until the supernatant was clear. Enterocytes were obtained after centrifugation at 1500 rpm for 5 min and resuspended in PBS. For the adhesion test, 106 enterocytes/mL were used.
Adhesion test
The adhesion test was performed according to the method of Nagayama et al. with modifications [14]. S. flexneri was cultured in BHI liquid for 24 h at 37 °C and then centrifuged at 6000 rpm at 4 °C for 10 min. The pellet was resuspended in PBS containing 1 % bovine serum albumin. The bacterial count was set at 108/mL using a spectrophotometer at a wavelength of 600 nm. Then, 0, 6.25, 12.5, 25, 50, or 100 µg of OMP was added to 300 µg of the enterocyte suspension and rocked gently on a shaking water bath at 37 °C for 30 min.
Finally, 300 µg of the bacterial suspension (108/mL) were added to each mixture, incubated on a shaking incubator for 30 min at 37 °C, and then centrifuged at 1500 rpm at 4 °C for 3 min. The pellet was washed twice with PBS. A sample was smeared on a glass slide and subjected to Gram staining.
Immunization of mice with S. flexneri OMP
The study used 6–8-week-old male BALB/C mice. The antigen used was the purified 28 kDa S. flexneri hemagglutinin adhesion OMP. Mice were acclimatized for 7 days before immunization. After acclimatization, the mice were injected intraperitoneally with 100 µg OMP that had been emulsified with 0.1 mL complete Freund’s adjuvant [18].
Isolation of polyclonal antibodies
Blood was collected from the hearts of the mice, placed in an Eppendorf tube, which was left tilted for 30 min, and then centrifuged at 10,000 rpm for 5 min. The supernatant serum was stored at –20 °C [8].
Antigen/antibody cross-reactions
The cross-reaction of Shigella spp. OMP antigen with antibody against the S. flexneri 28 kDa OMP was examined by Western blotting. Before performing the antigen antibody response test, the checkerboard test was necessary to determine the effectiveness of the antigen and antibody reactions to be evaluated. The checkerboard test uses a dot blot method and reacts the S. flexneri 28 kDa OMP antigen with serially diluted antibodies. The antigen was diluted 1/5 to 1/10240, and the antibody was diluted 1/100 to 1/2800. The results reveal a color gradation that can be analyzed using Corel Photo-Paint to determine the mean value [19]. Western blotting was performed using the method of Towbin et al [20].
Statistical analysis
The data were analyzed using SPSS 23. One-way analysis of variance with Tukey’s post hoc test was used to determine significant differences in the mean index of adhesion among the sample groups. Pearson’s correlation test was used to determine correlations between the sample doses and adhesion index. Significance level was set at p ˂ 0.05.
Results
OMP Shigella spp profiling
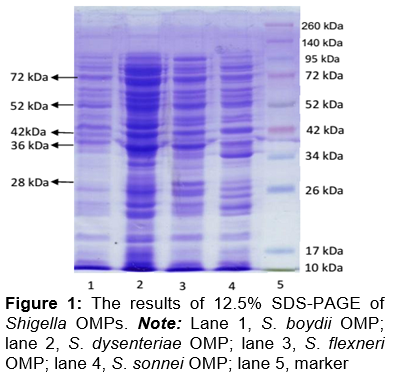
shows the molecular weights of the four Shigella species OMPs determined using SDS-PAGE. The profiles of the Shigella spp. OMPs were identical. There were various Shigella spp. OMPs, including OMPX, OMPA, OMPT, OMPLA, TOLC, α-hemolysin, porin OMP32, FhuA, and FEPA [5].
Hemagglutination
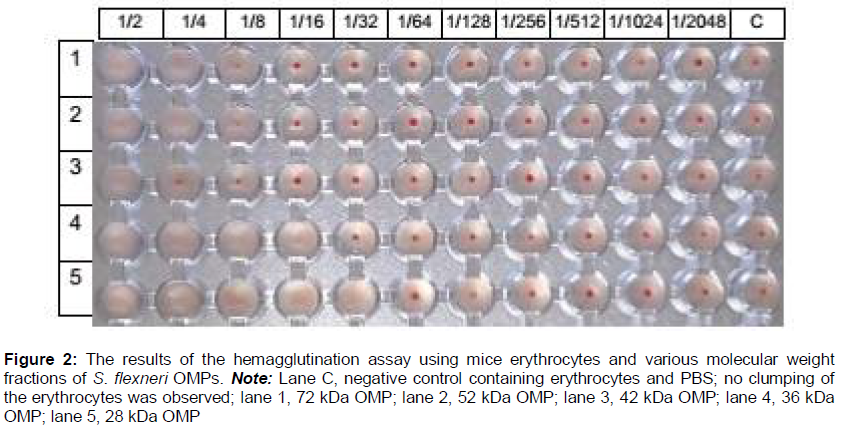
The S. flexneri OMP bands with molecular weights of 28, 36, 42, 52, and 72 kDa were identified as candidate adhesion proteins. These five protein bands were excised, collected, and purified by electro elution and dialysis for the hemagglutination test. The hemagglutination test results shown in indicated that the 28 kDa OMP had the highest hemagglutination titer (1/32), and thus it was selected for the adhesion tests.
Adhesion
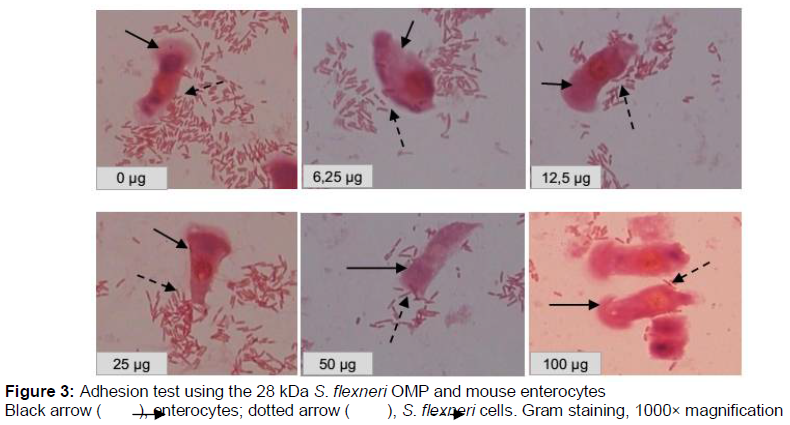
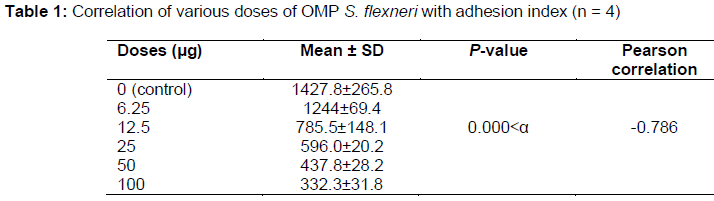
The adhesion test was conducted using 0 (control), 6.25, 12.5, 25, 50, and 100 µg of the 28 kDa S. flexneri OMP (). The adhesion index was calculated by counting the number of bacteria attached to 100 enterocytes. The result is shown in , which shows that the number of S. flexneri attached to mouse enterocytes decreased with increasing OMP dose. There was a significant and strong negative correlation between the dose and adhesion index (p=0.000). This proved that OMP inhibited the attachment of S. flexneri to enterocytes.
Antigen/antibody cross-reaction
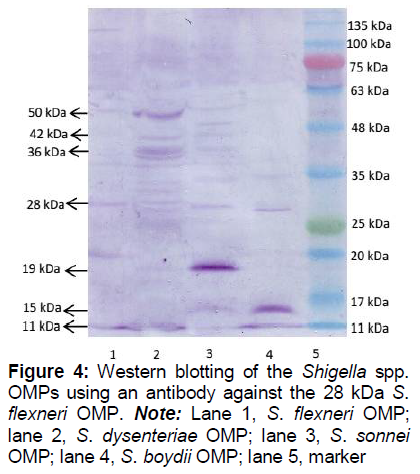
In the checkerboard test, the S. flexneri OMP antigen diluted 1/320 and antibodies against the 28 kDa S. flexneri OMP diluted 1/400 showed the lowest density (data not shown). In this zone, there is a balanced antigen to antibody ratio. In Western blotting, antibodies against the 28 kDa S. flexneri OMP diluted 1/400 reacted with Shigella spp. OMP.
In , lane 1 represents the 28 kDa S. flexneri OMP reacting with the antibody against the 28 kDa S. flexneri OMP, demonstrating that the 28 kDa S. flexneri OMP is an antigen capable of stimulating an antibody response. OMPs from other Shigella spp. (, lanes 2 to 4) also reacted with the 28 kDa S. flexneri OMP antibody.
Discussion
Shigellosis is caused by four Shigella species: S. flexneri, S. dysenteriae, S. boydii, and S. sonnei [4]. Shigella spp. OMP was isolated after shaving the bacteria pili using a pili bacterial cutter. The profiles of the Shigella spp. OMPs on SDS-PAGE gels were identical, and their calculated molecular weights were similar. The number of OMP bands observed on SDS-PAGE gels indicated that Shigella species expressed many OMPs of various molecular weights, including OMPX, OMPA, OMPT, OMPLA, TOLC, α-hemolysin, porin OMP32, FhuA, and FEPA [5].
The S. flexneri 28 kDa OMP had the highest hemagglutination titer (1/32). Hemagglutination is an indicator of the ability of bacteria to attach to animal cells, because the erythrocyte membrane receptor is homologous to the mucosal host cell surface receptors [21].In order to confirm that the S. flexneri 28 kDa OMP is an adhesion protein, we conducted adhesion tests. The results showed that the adhesion of S. flexneri to mouse enterocytes was inhibited by the S. flexneri 28 kDa OMP, as shown in and . There was a significant (p=0.000) correlation between the dose and adhesion index. Therefore, OMP was able to inhibit the attachment of S. flexneri to enterocytes. We conclude that the 28 kDa S. flexneri OMP was an adhesin protein and a bacterial virulence factor [10]. Histopathological studies have demon-strated that inflammatory lesions in shigellosis are caused by the entry of bacteria into the upper rectum and distal colon. [4]. Immunization of mice with the 49.8 kDa S. dysenteriae pili adhesion protein resulted in minimal colonic epithelial cell damage with elevated s-IgA levels. s-IgA binds to intracellular enterocyte antigens, minimizing the destruction of epithelial cells in the intestinal mucosa [9]. The 49.8 kDa S. dysenteriae pili adhesion protein also protected against the movement of solution from the intestine to lumen in the MLIL model [14].
The antigen–antibody cross-reaction examined by Western blotting showed that many Shigella spp. OMPs reacted with the antibody against the S. flexneri 28 kDa OMP (). Therefore, the OMP antibody would be capable of recognizing epitopes present in the OMPs of all Shigella species. Antibodies produced against one antigen may bind to a different structurally related antigen, referred to as a cross-reaction [22].These results are consistent with another research who detected cross-reaction between a 34 kDa OMP and intact cells of Shigella species. An antibody against the 34 kDa S. flexneri OMP recognized an epitope on the surface of S. flexneri [23]. After identifying the antigen–antibody response to Shigella OMPs, we postulated that the antibody would also prevent the adhesion of Shigella to the host cell, and hence could serve as a candidate shigellosis vaccine against all Shigella species.
The 34 kDa S. flexneri OMP has already been purified with a promising subunit vaccine candidate for shigellosis [11, 23]. Our hemagglutinin assay also showed a positive result for a 36 kDa S. flexneri OMP, which may also be an adhesion molecule ().Mitra et al found that the colonization ability of Shigella spp. was directly correlated with the hemagglutination activity in a suckling mouse model [24]. A recent in vivo study by Hamong et al. showed that a 49.6 kDa Helicobacter pylori hemagglutinin protein prevented damage to the stomach caused by homologous bacteria in mice [25].
We postulated that adhesion molecule epitopes are involved in protein hemagglutinin due to various OMPs [14]. This finding should be clarified before developing peptide vaccines based on molecule adhesion epitopes.
Conclusion
A 28 kDa S. flexneri OMP is identified here in this study as a hemagglutinin protein that serves as an adhesion protein. Antibodies against this protein cross-react with the OMPs of other Shigella species. Further research is needed to determine whether the 28 kDa S. flexneri OMP hemagglutinin adhesion protein can protect against colon epithelial cell damage and the movement of solution into the intestinal lumen in MLIL model [10].
Declarations
Acknowledgement
References
Archives


News Updates